Download to read offline

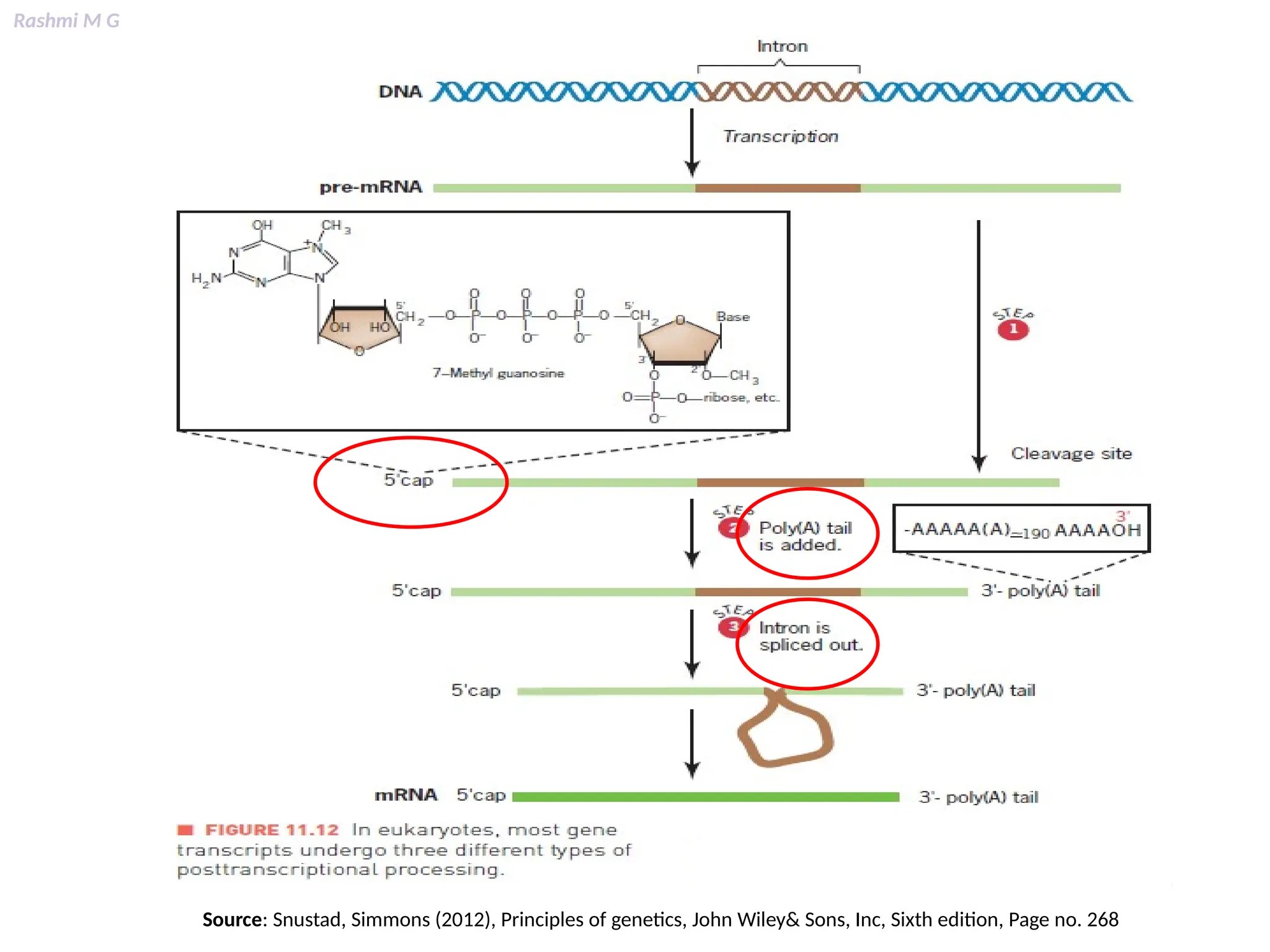
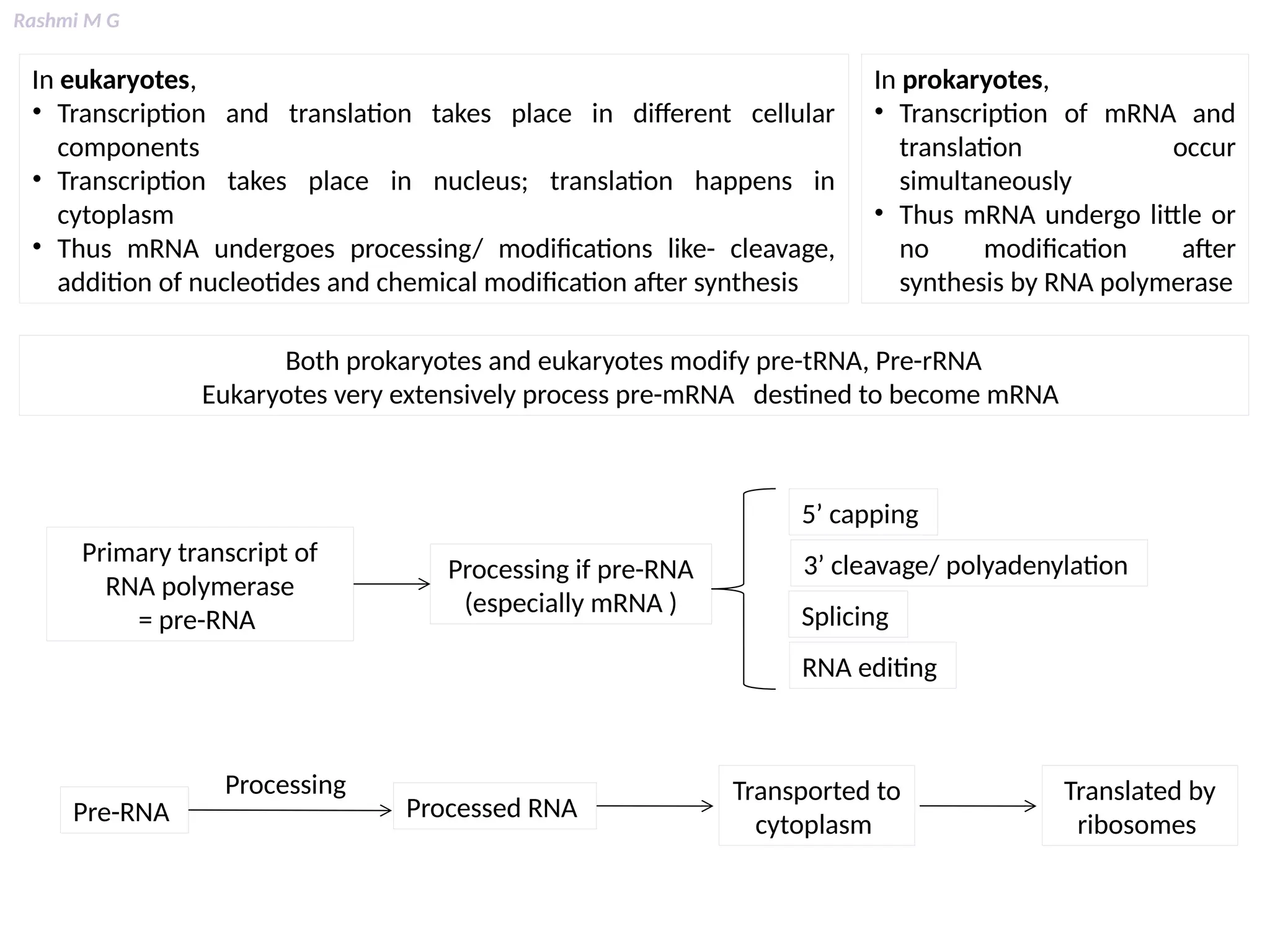
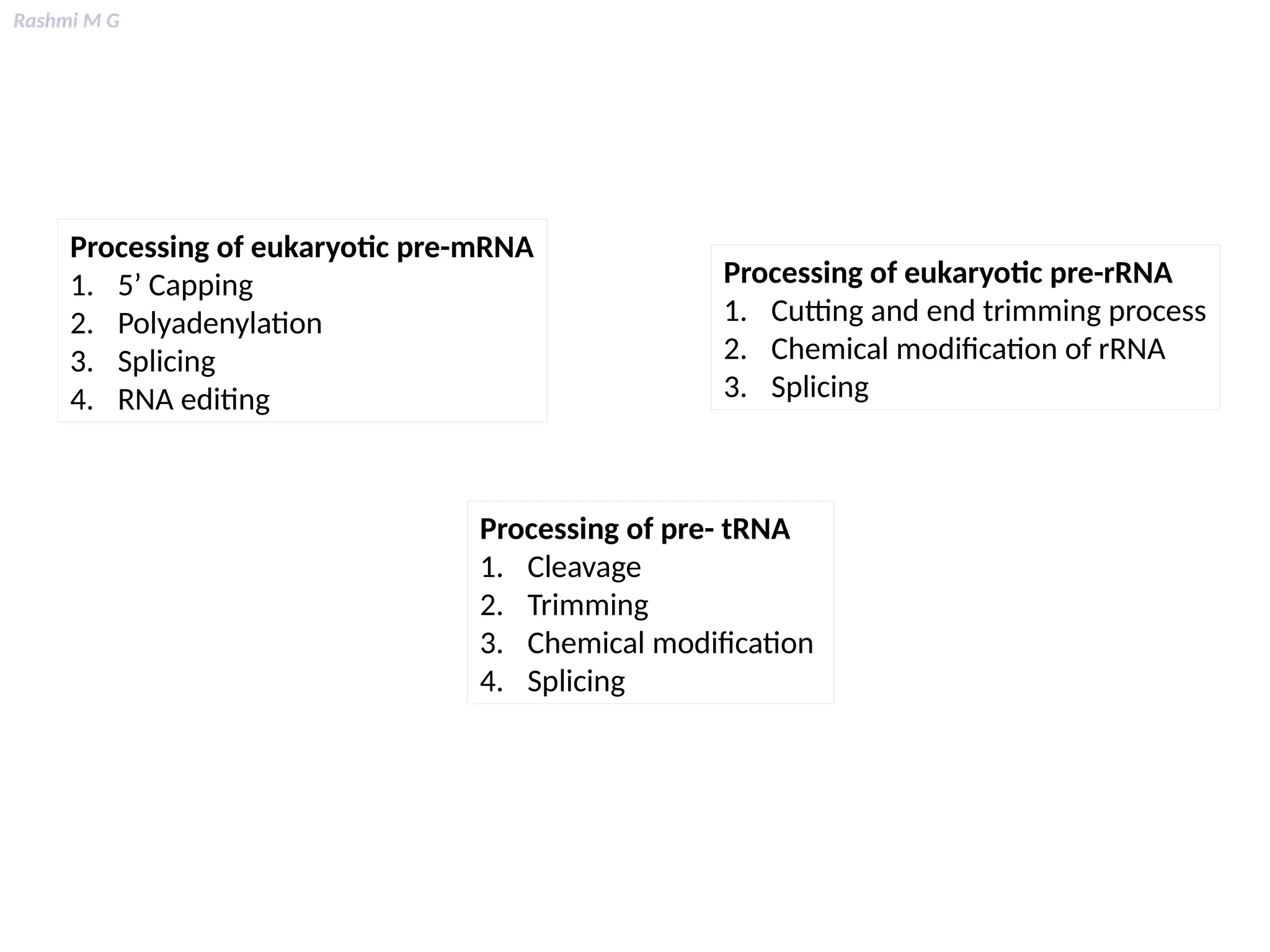
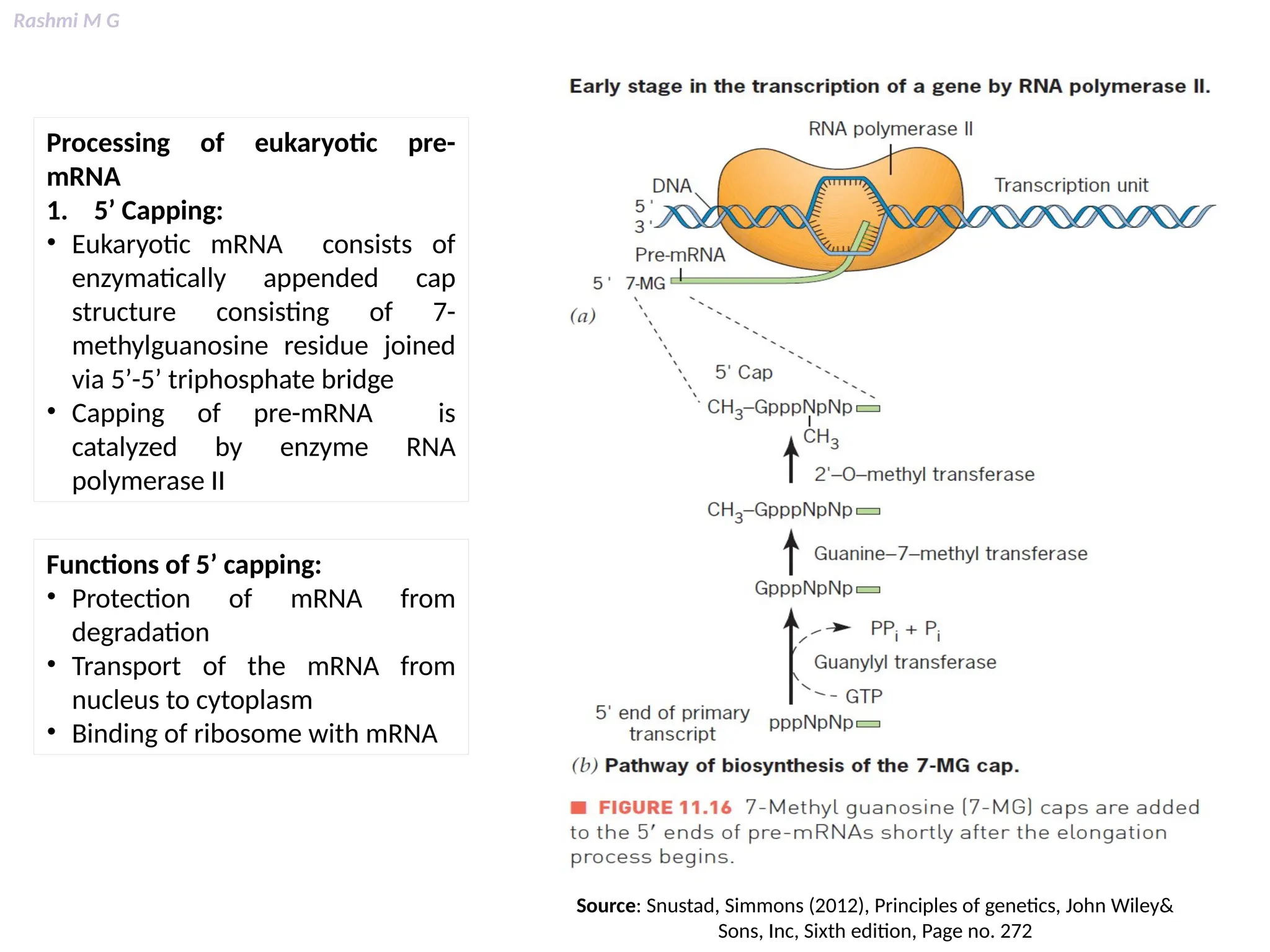
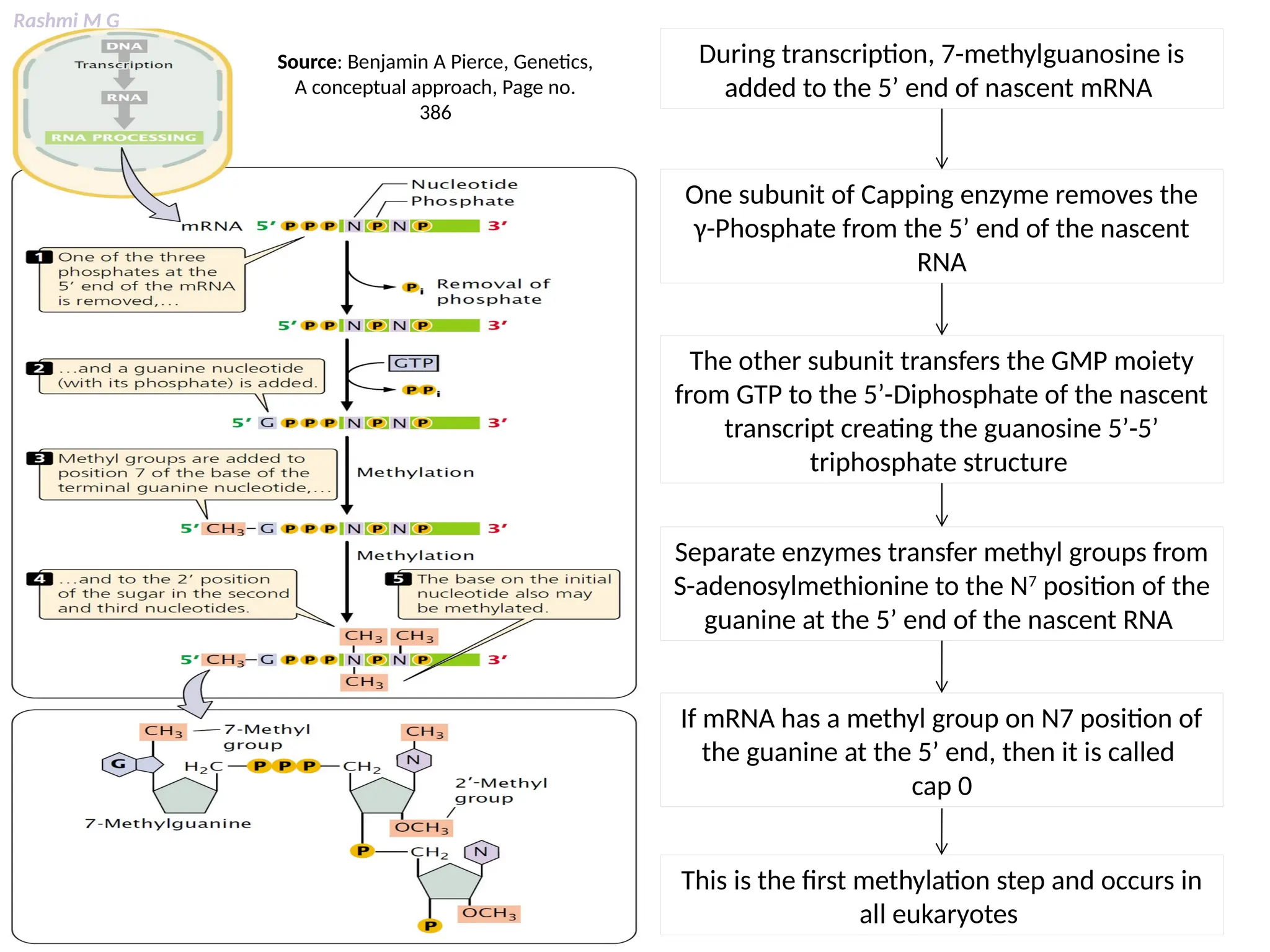

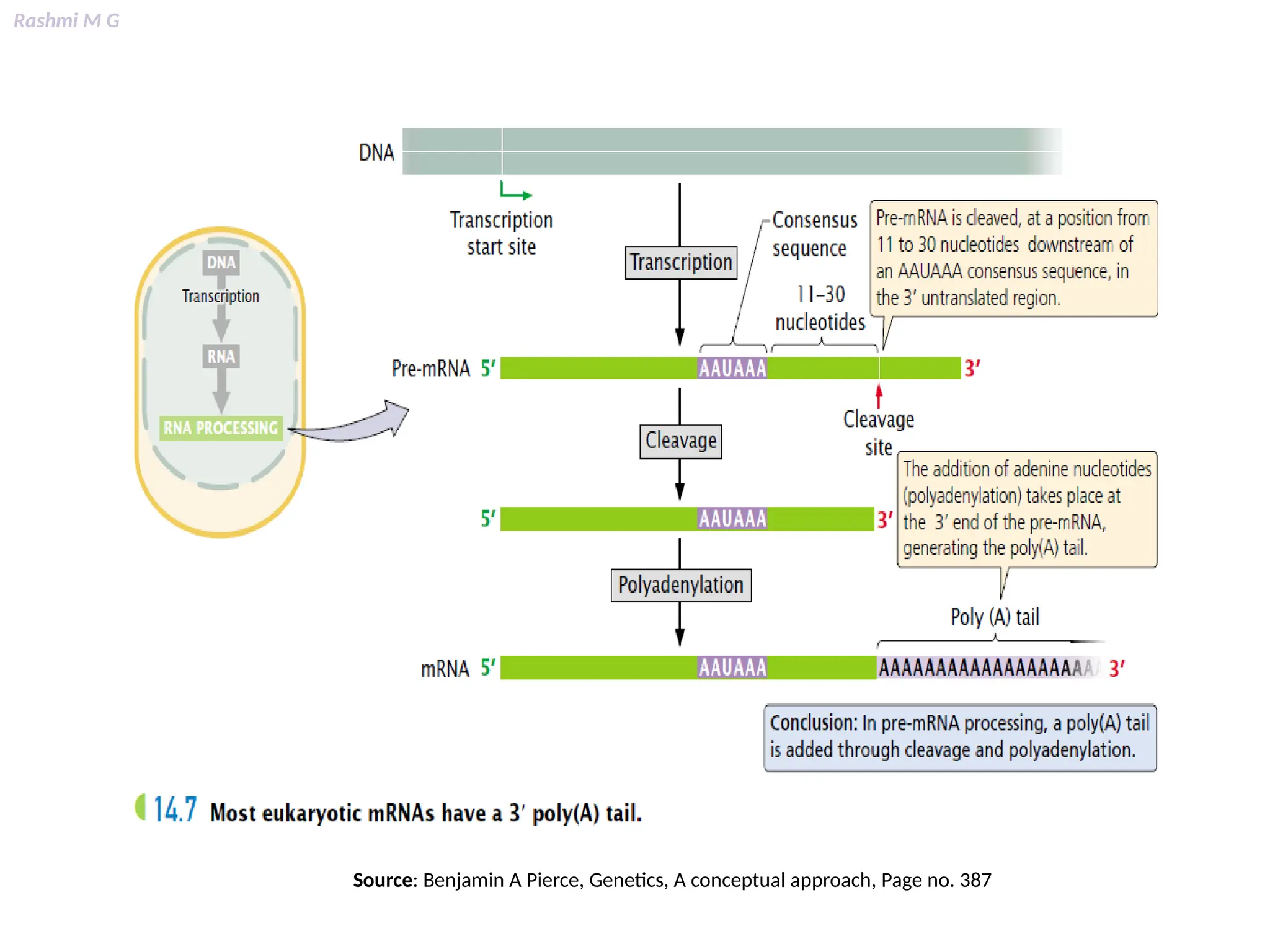



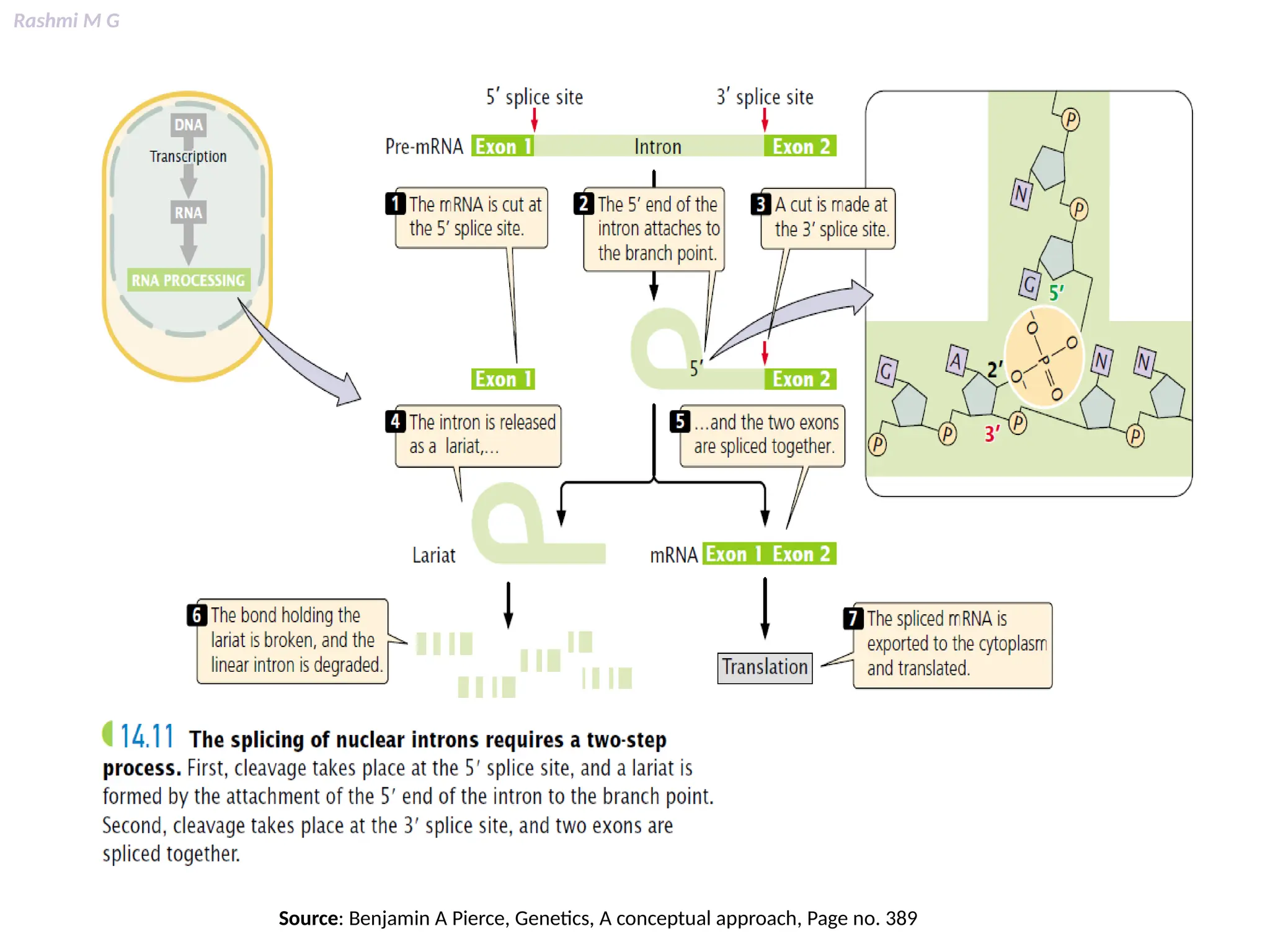


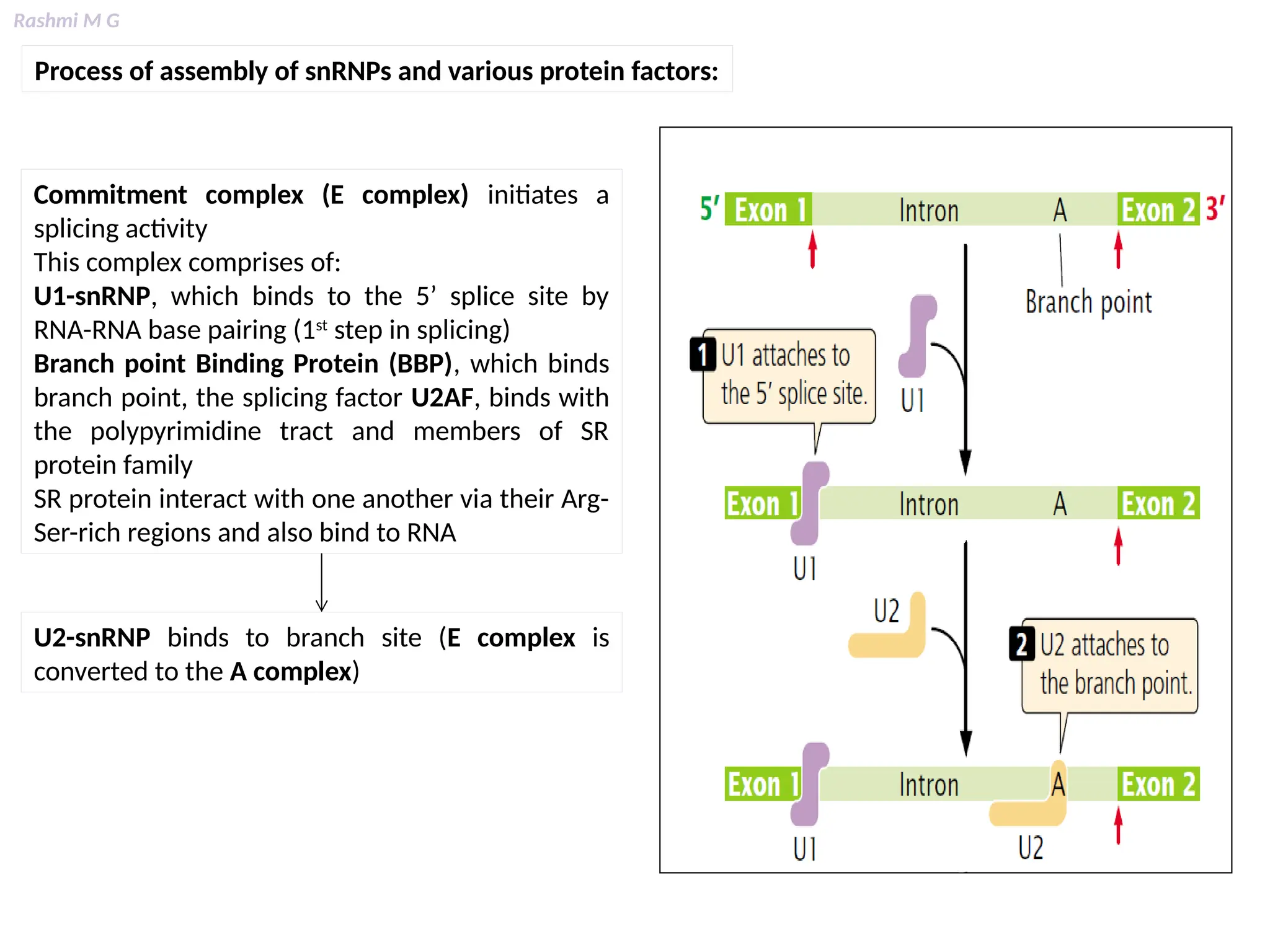
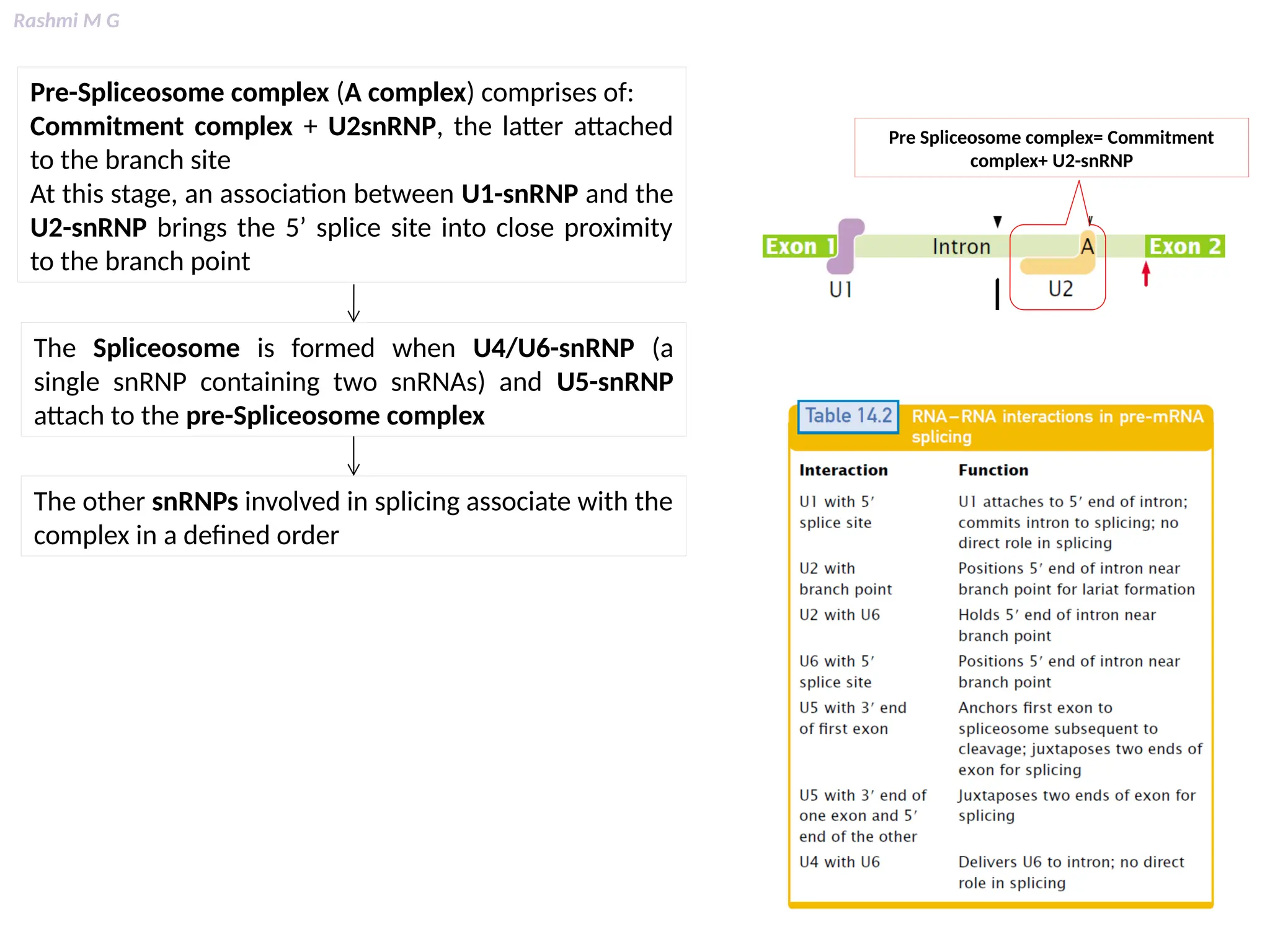
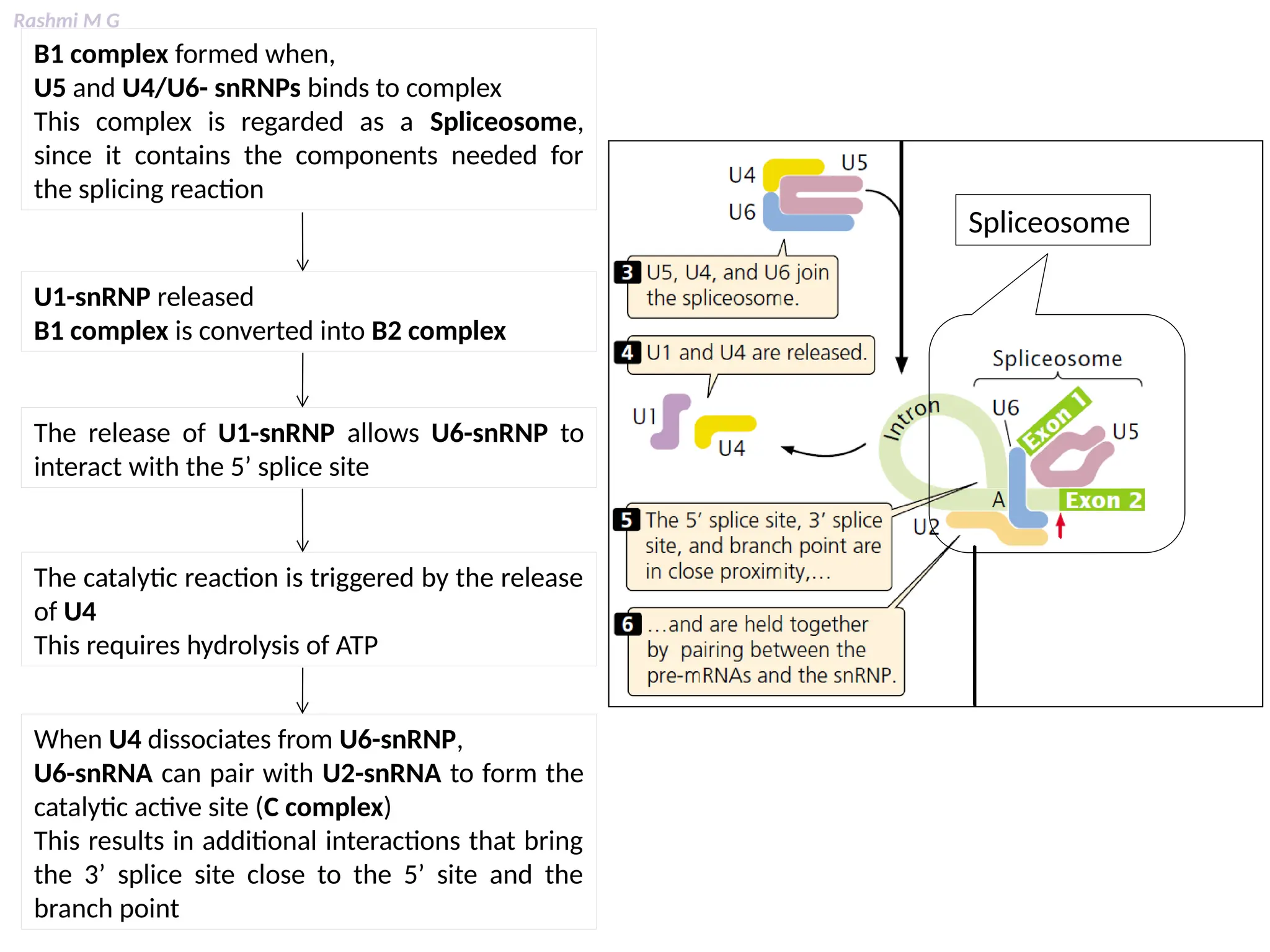
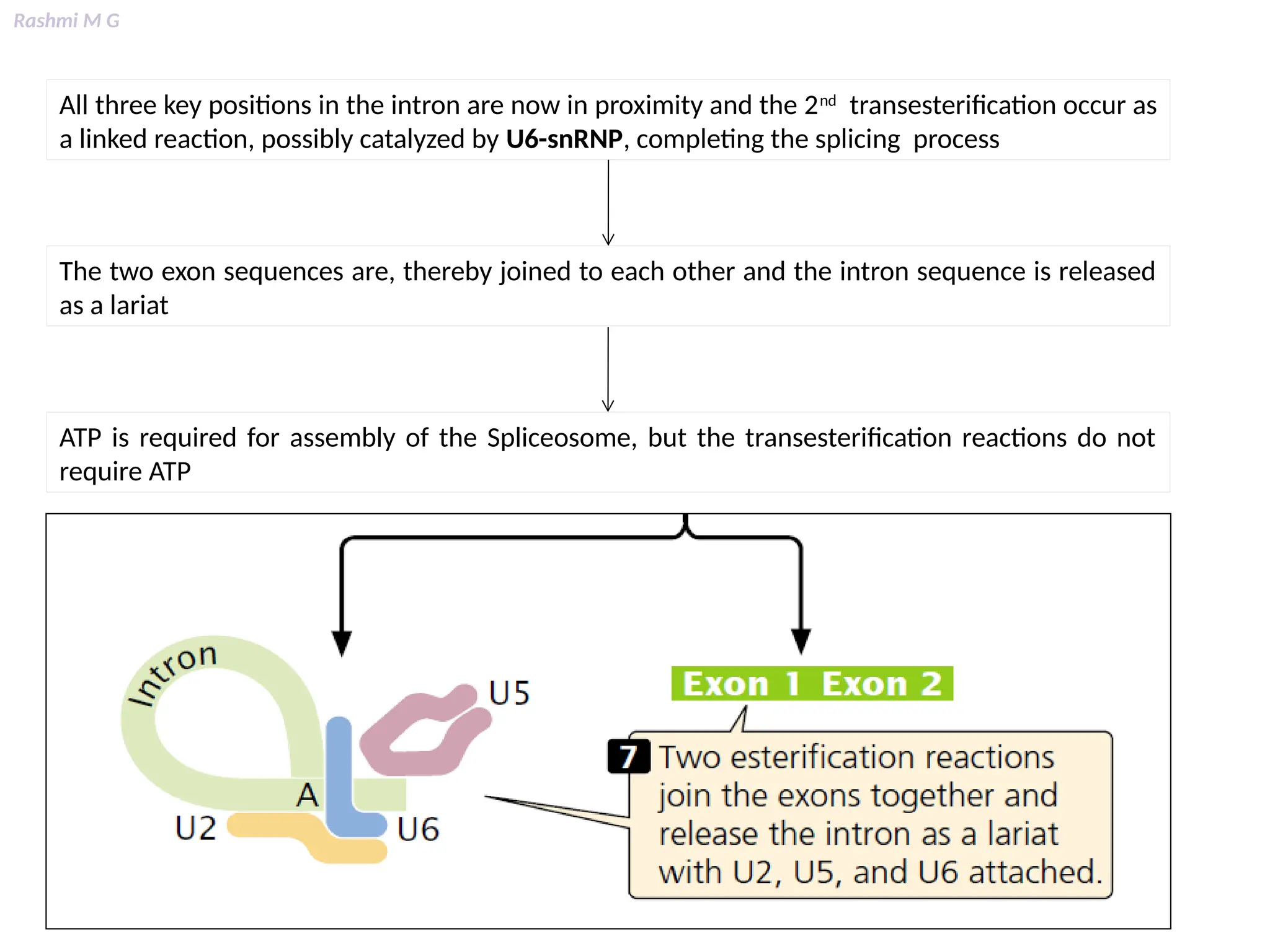
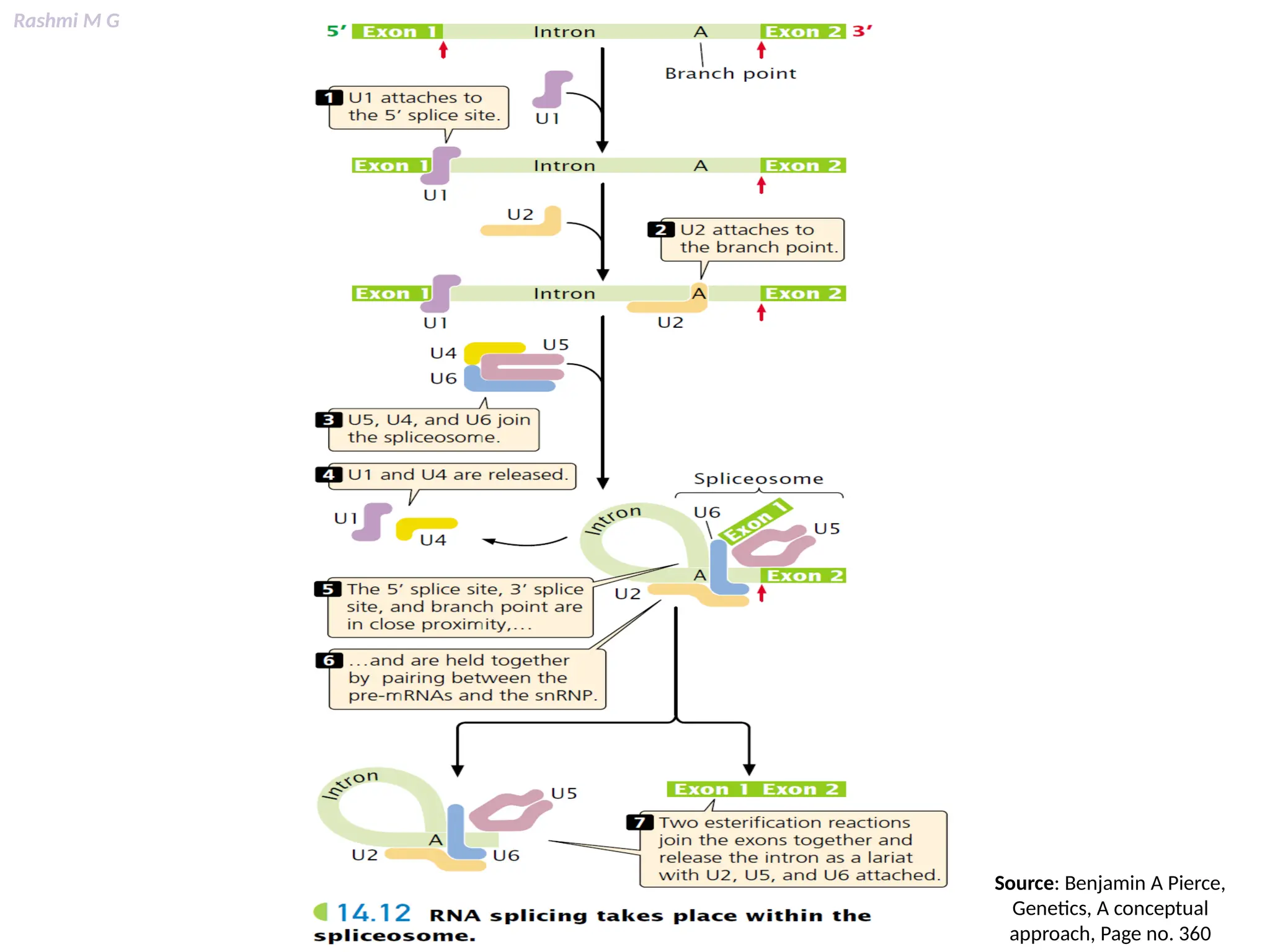

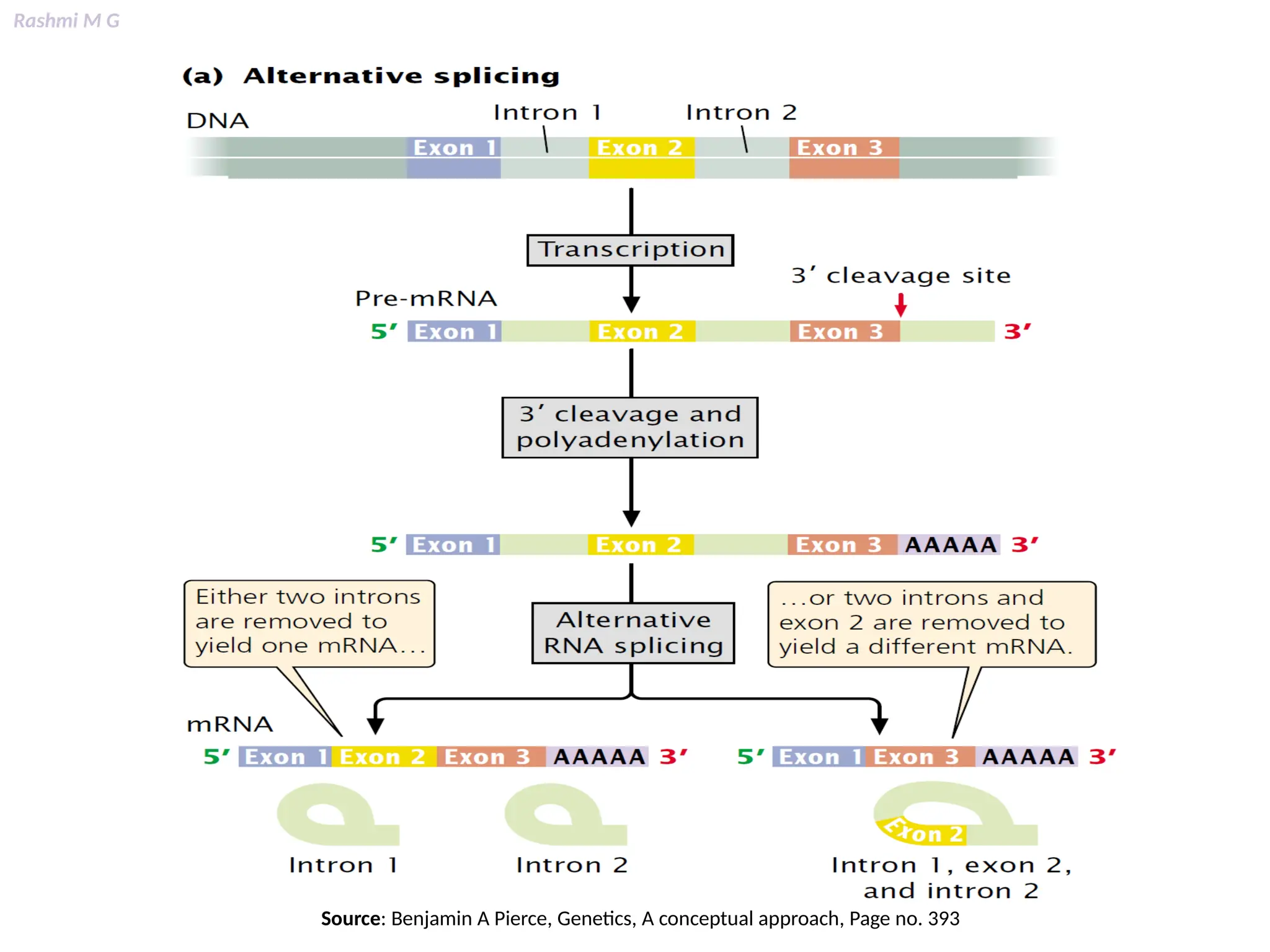
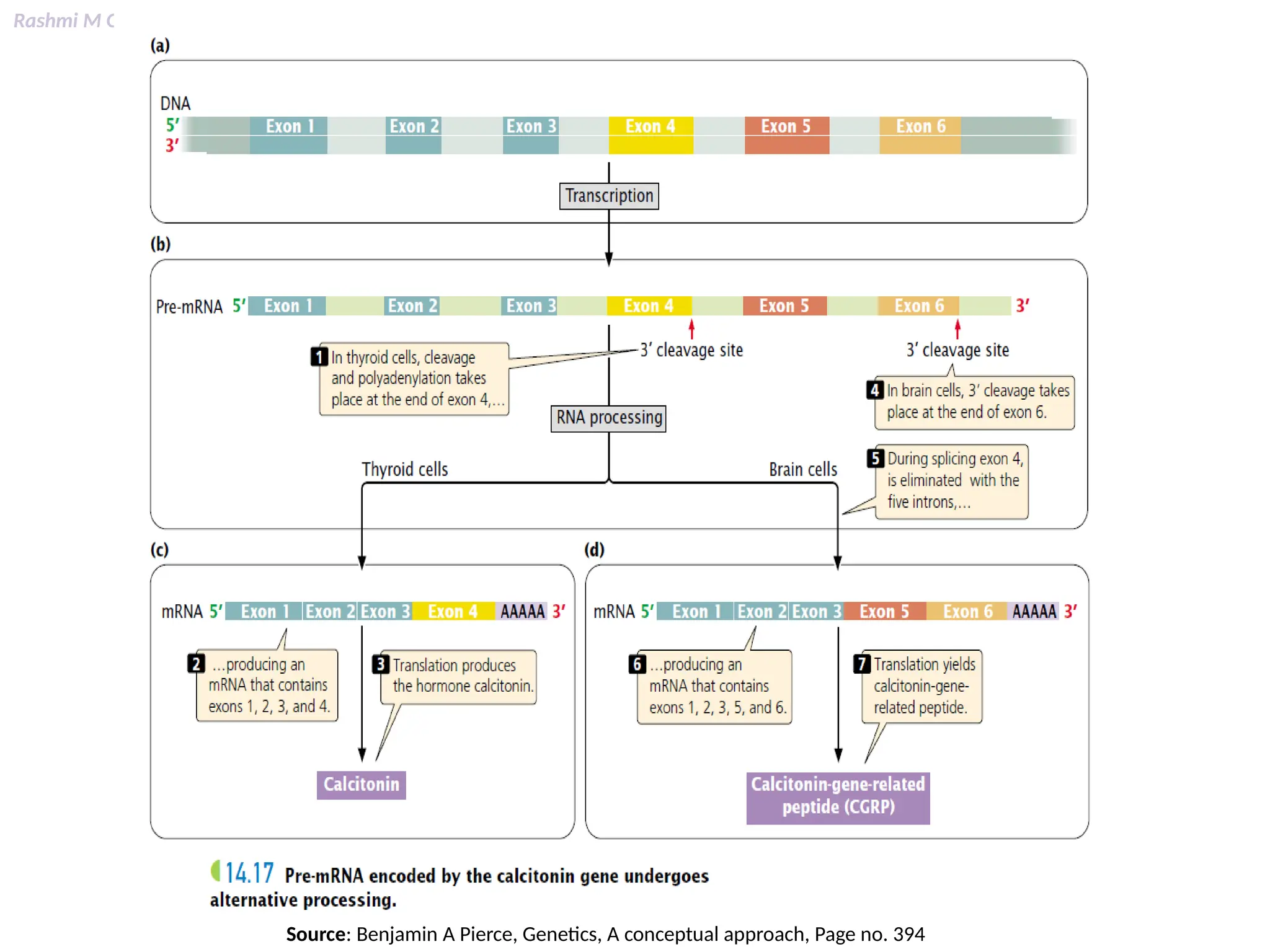
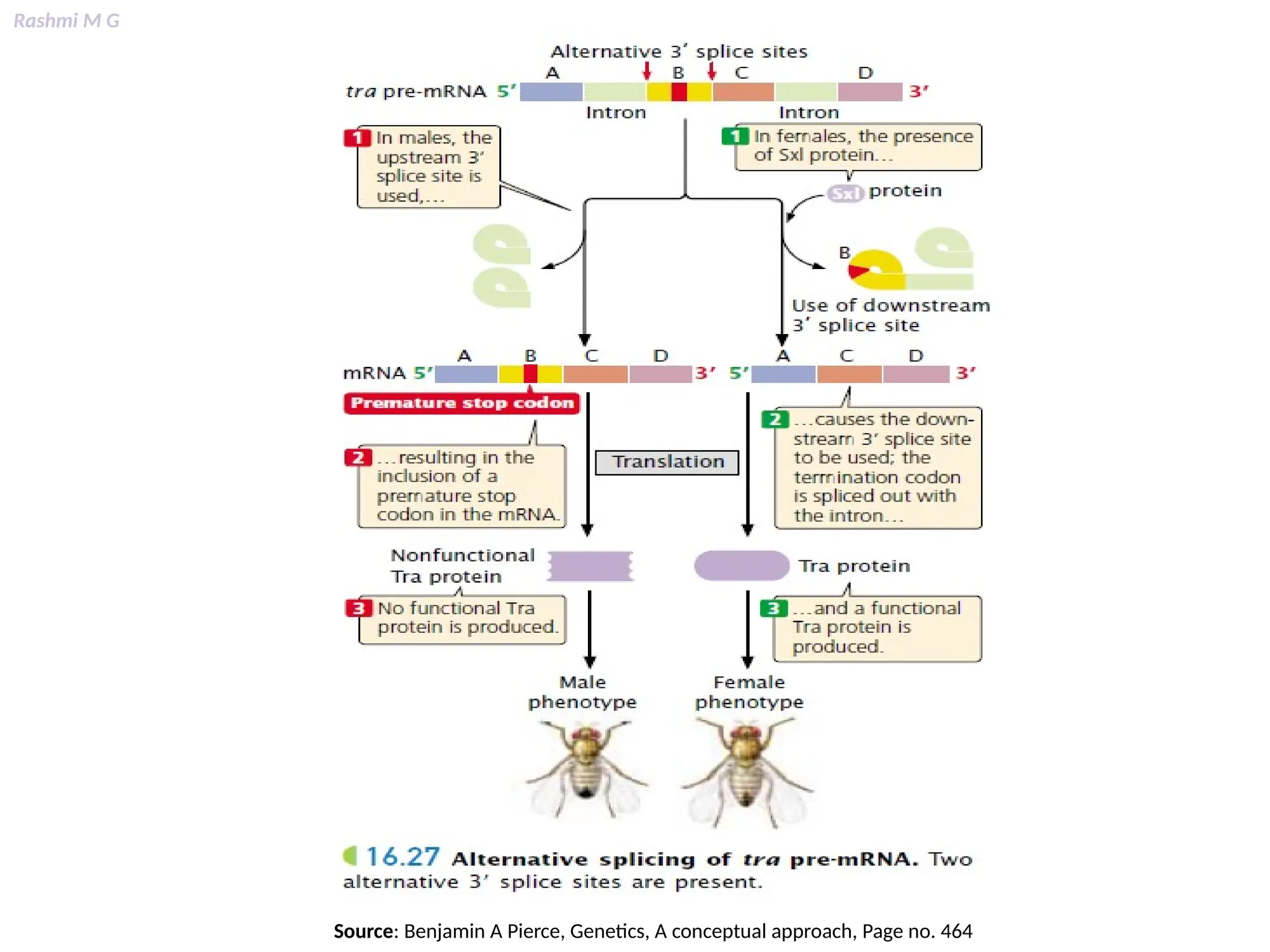
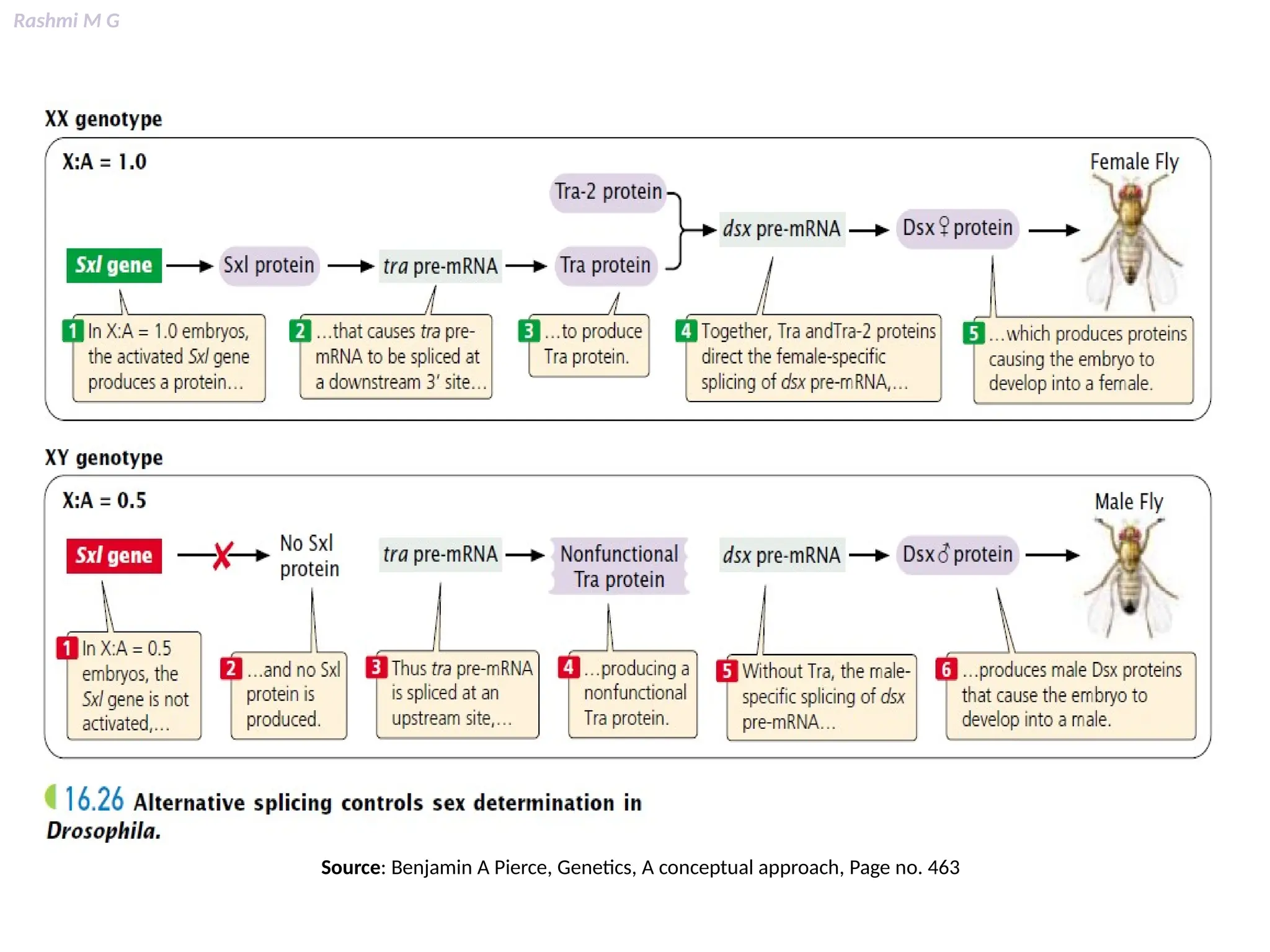
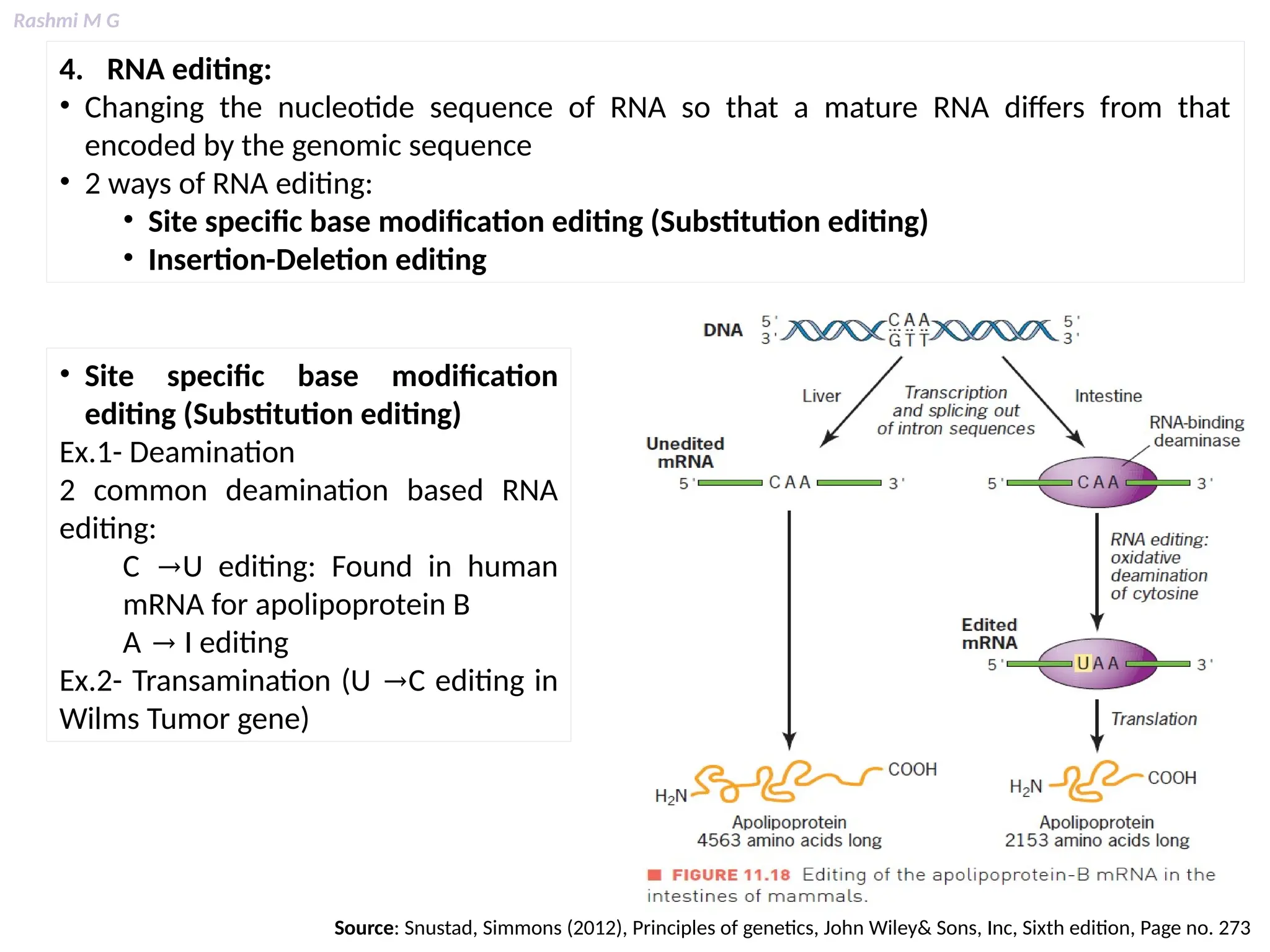

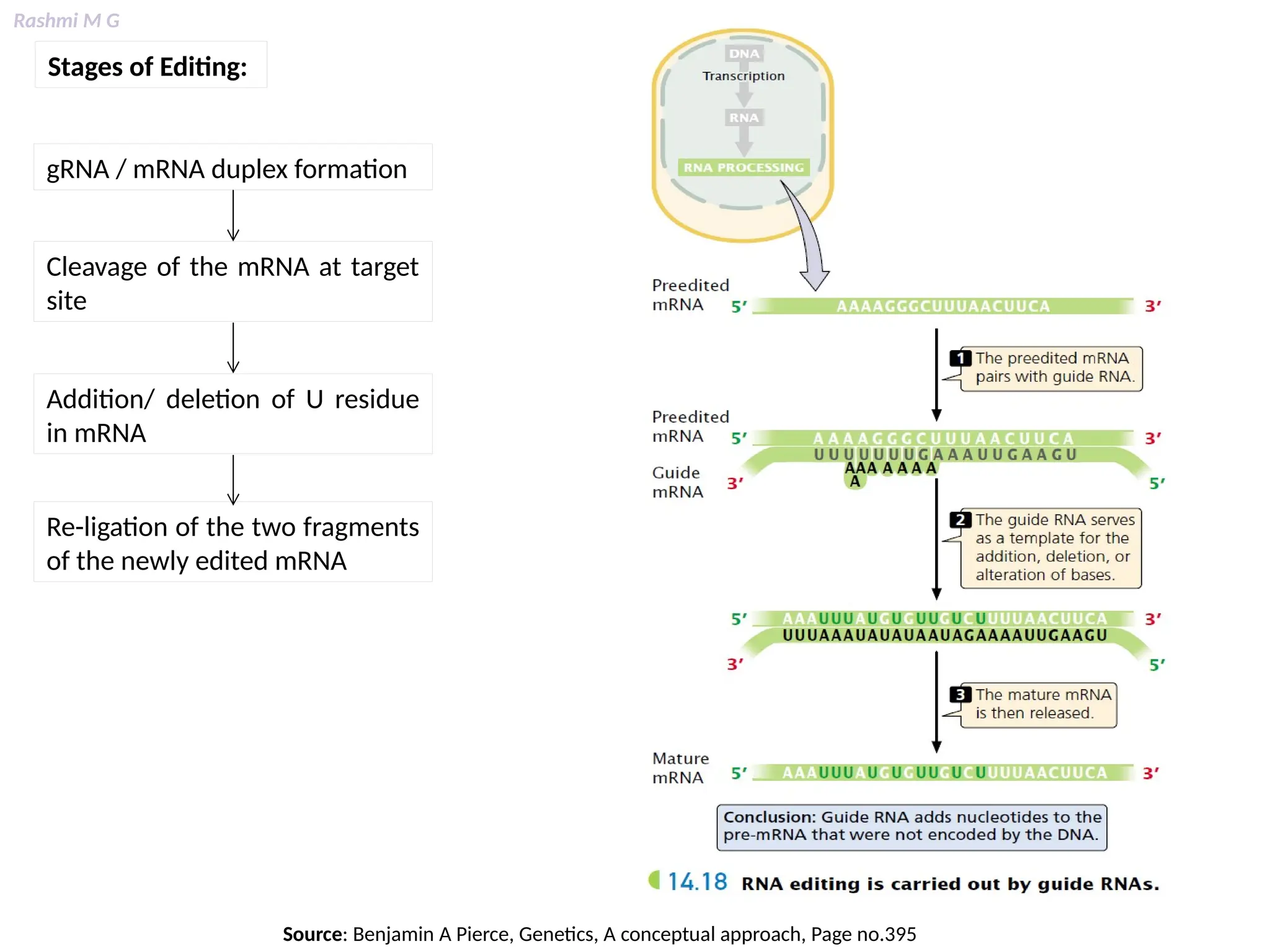

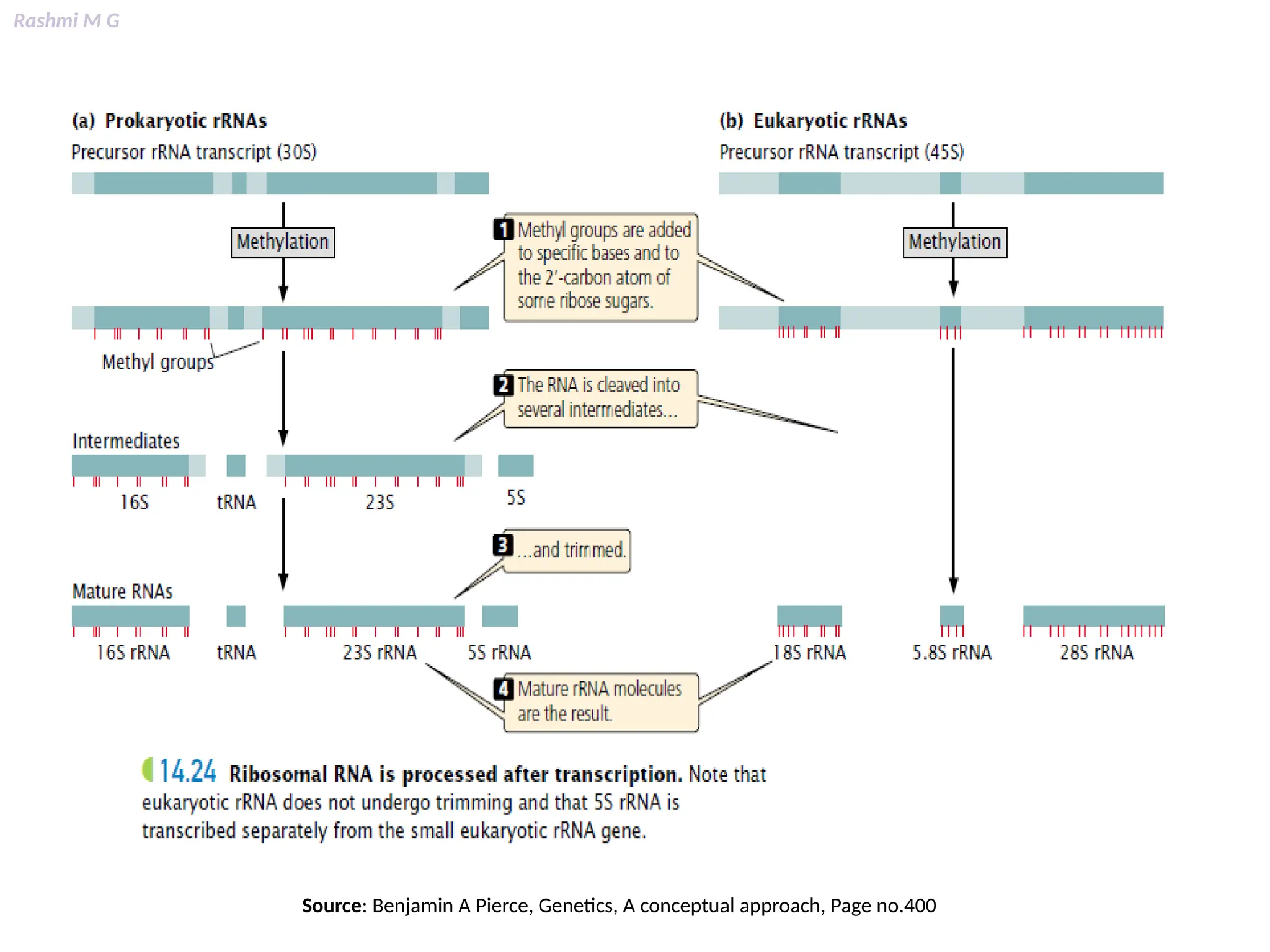


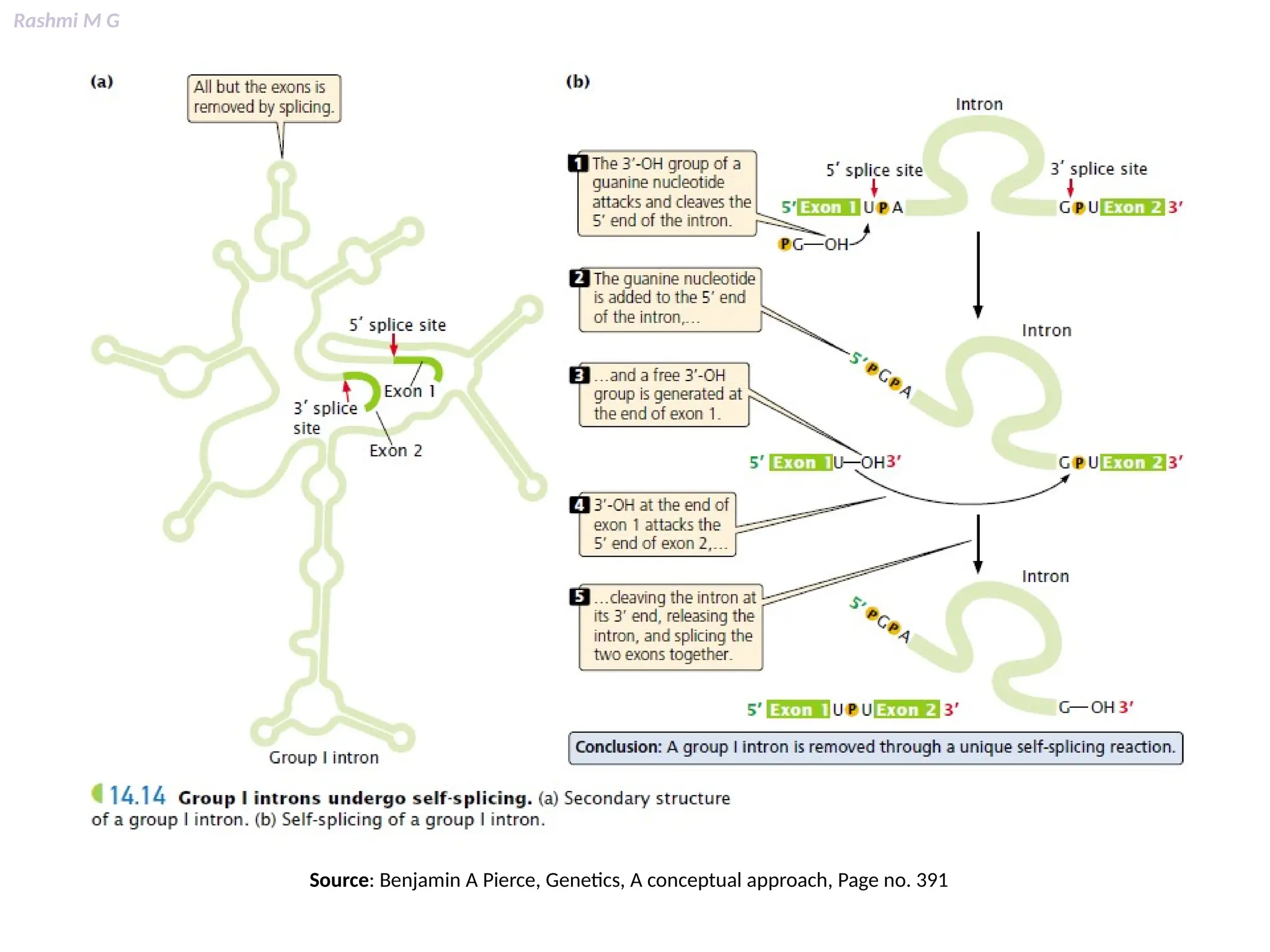
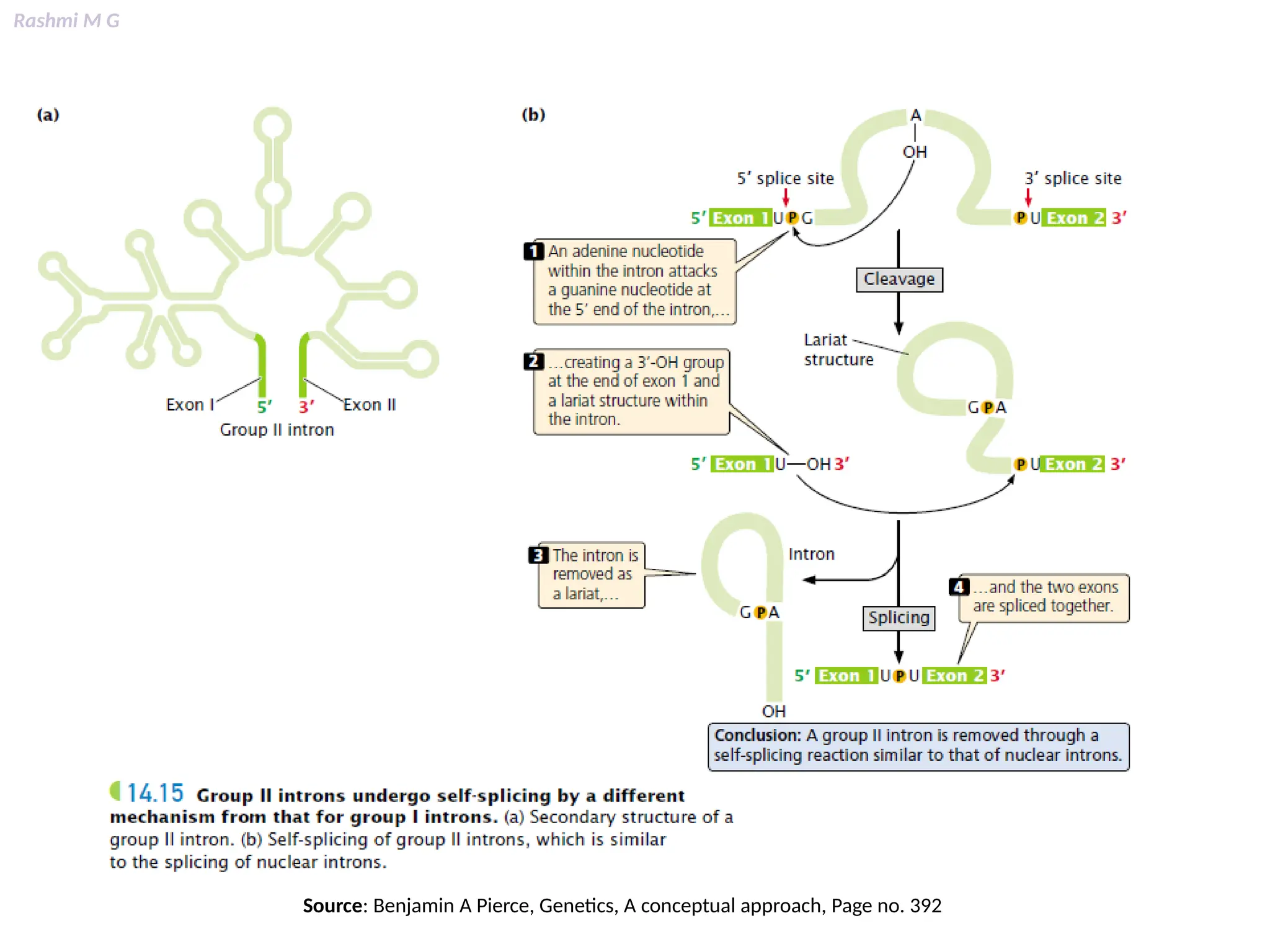
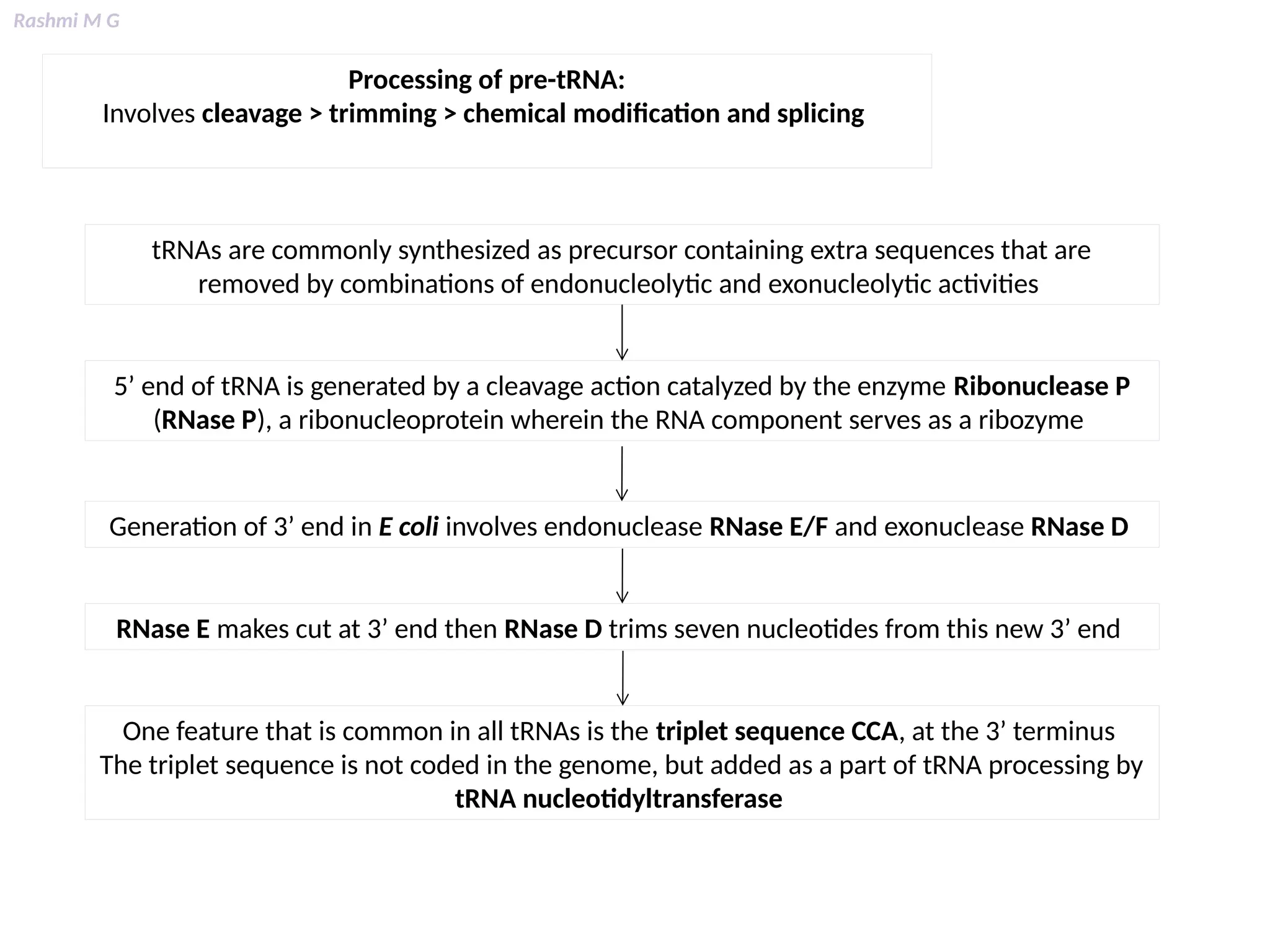
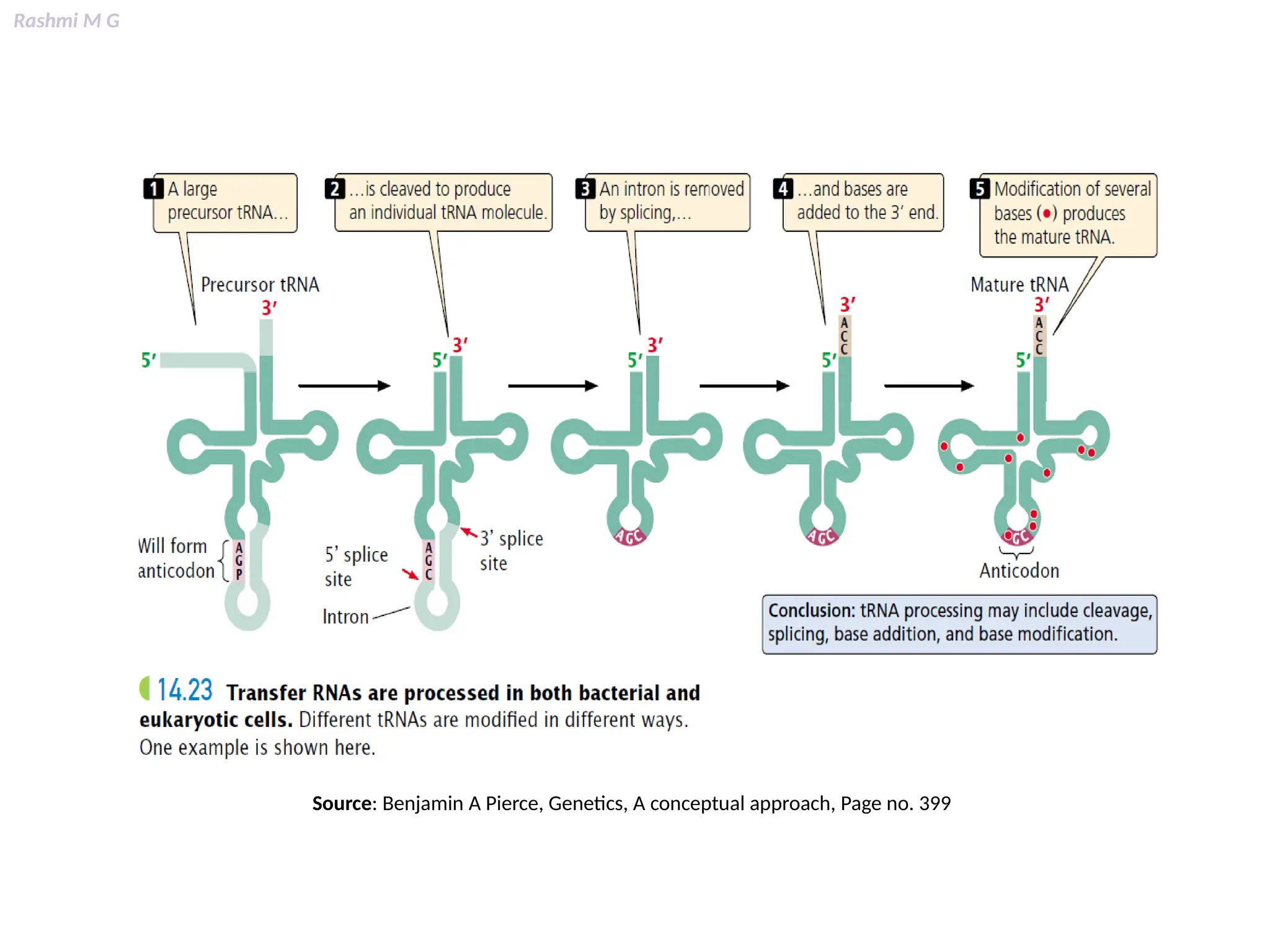

















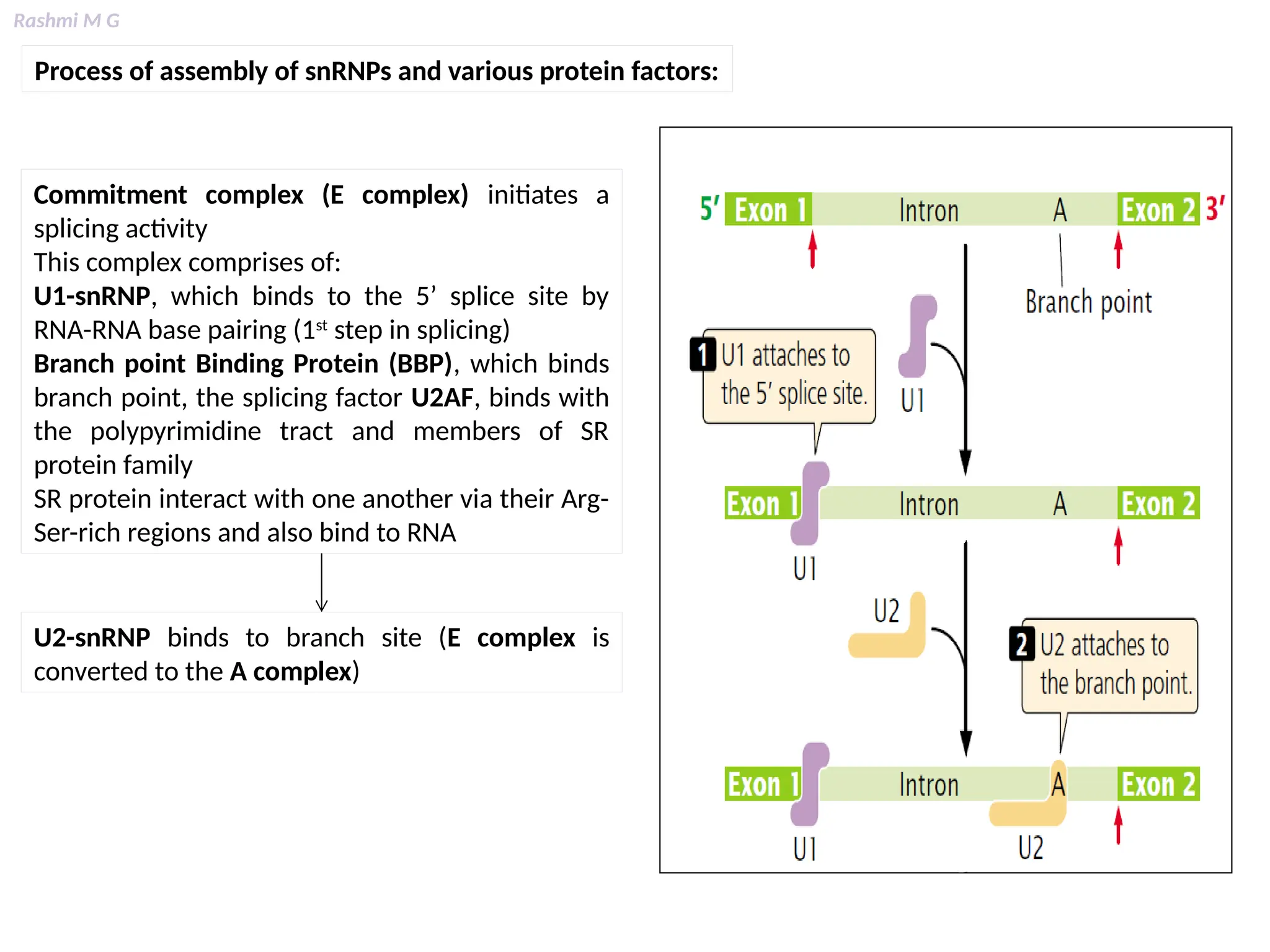

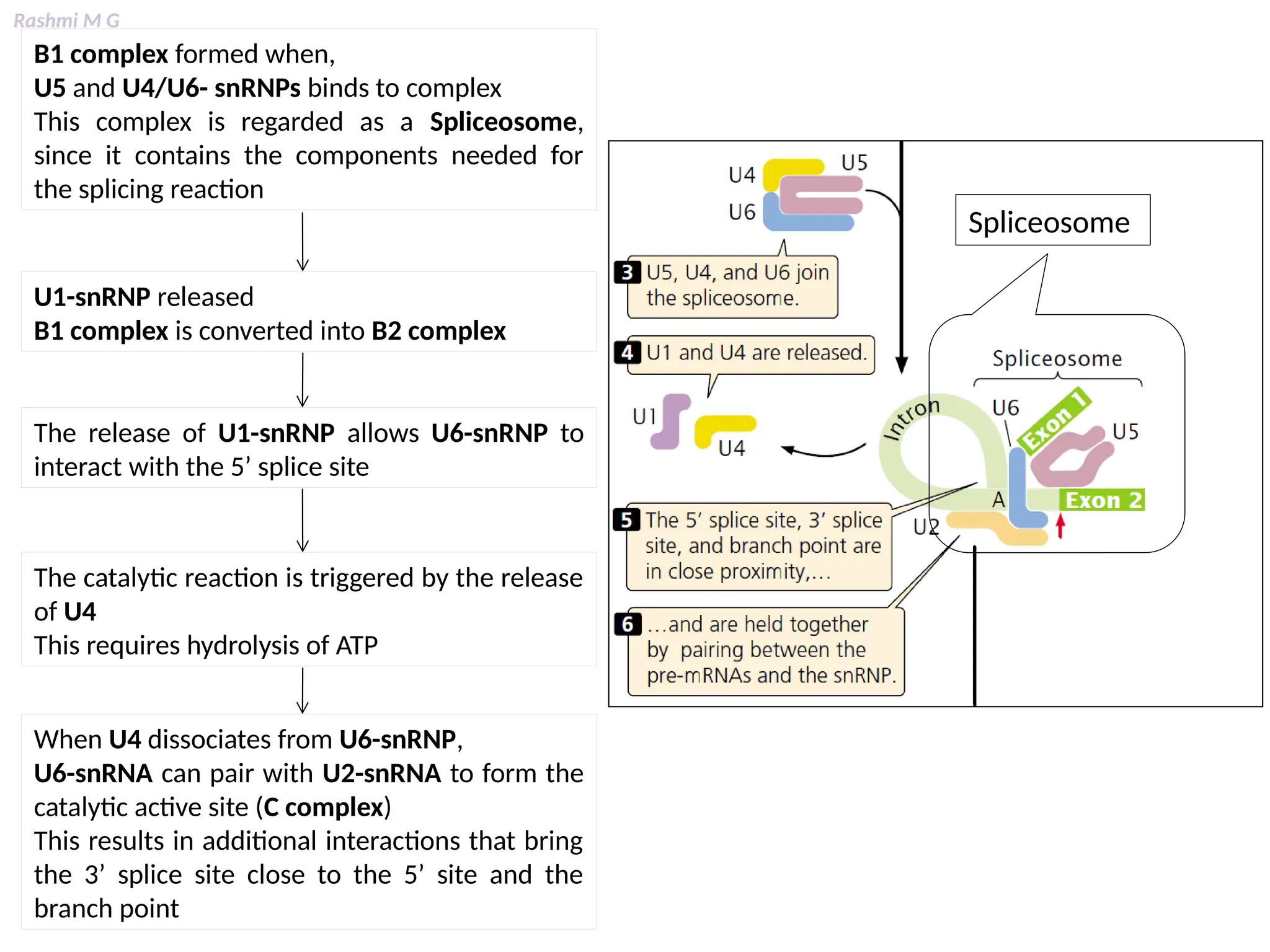
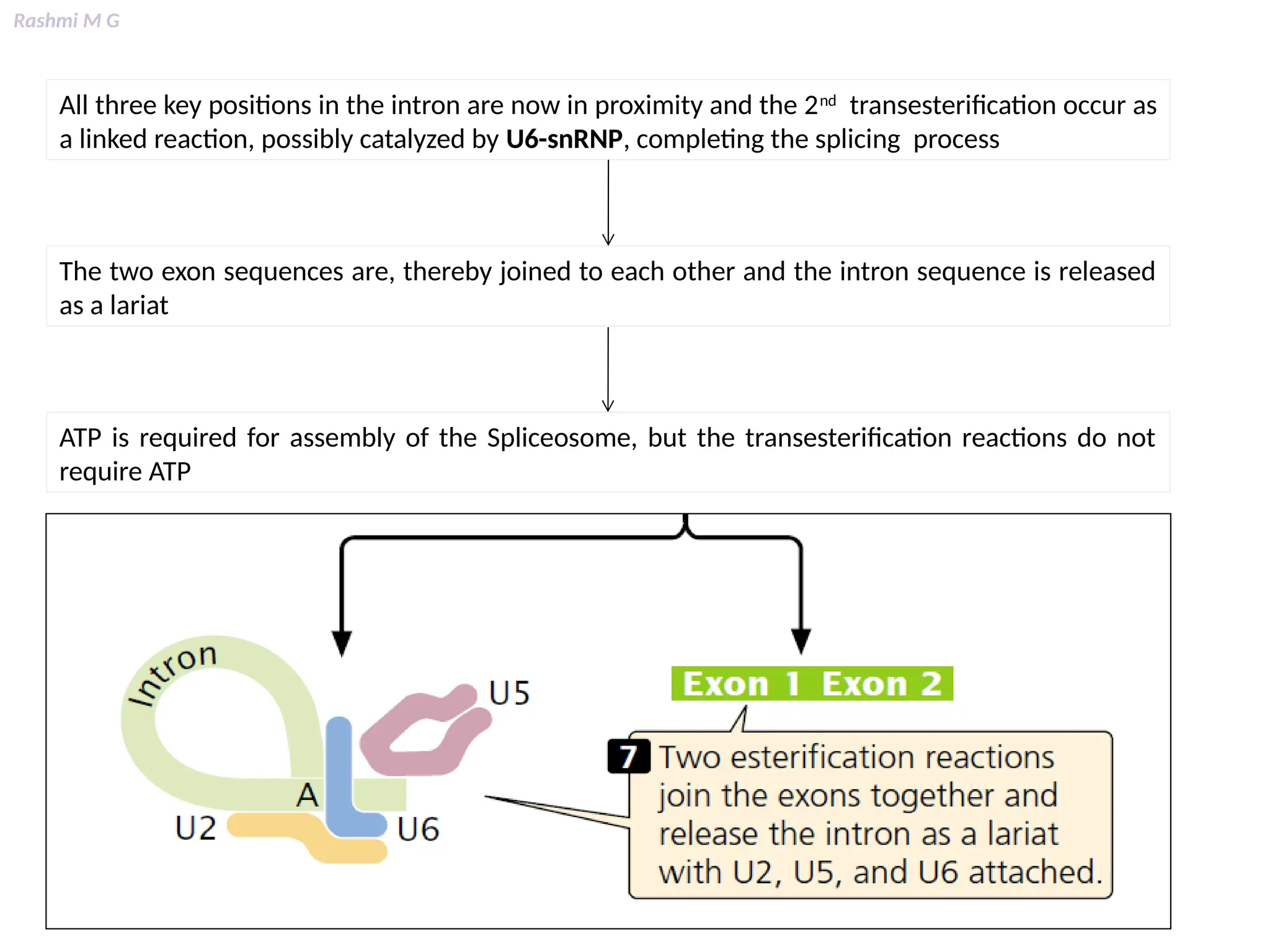








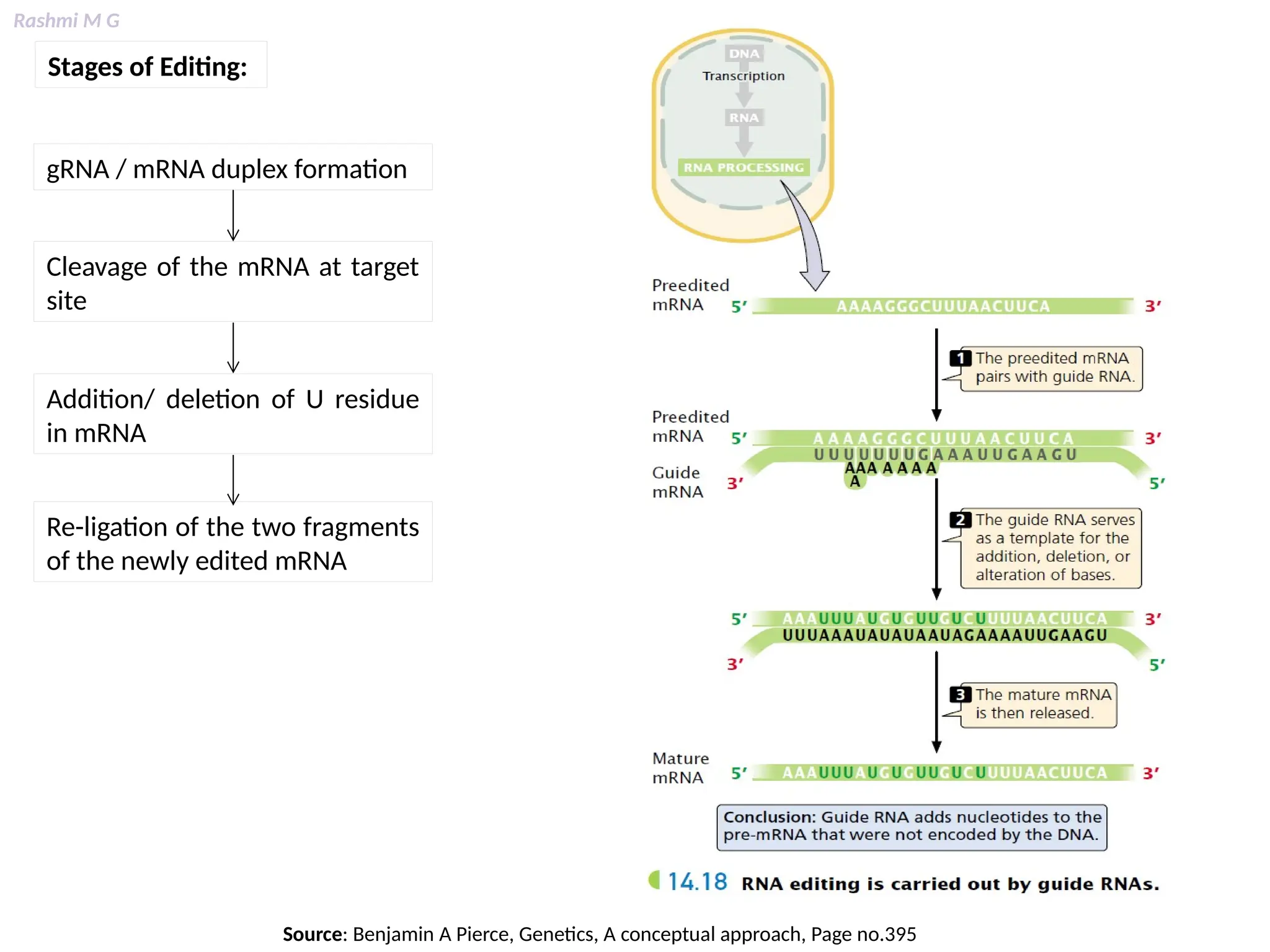




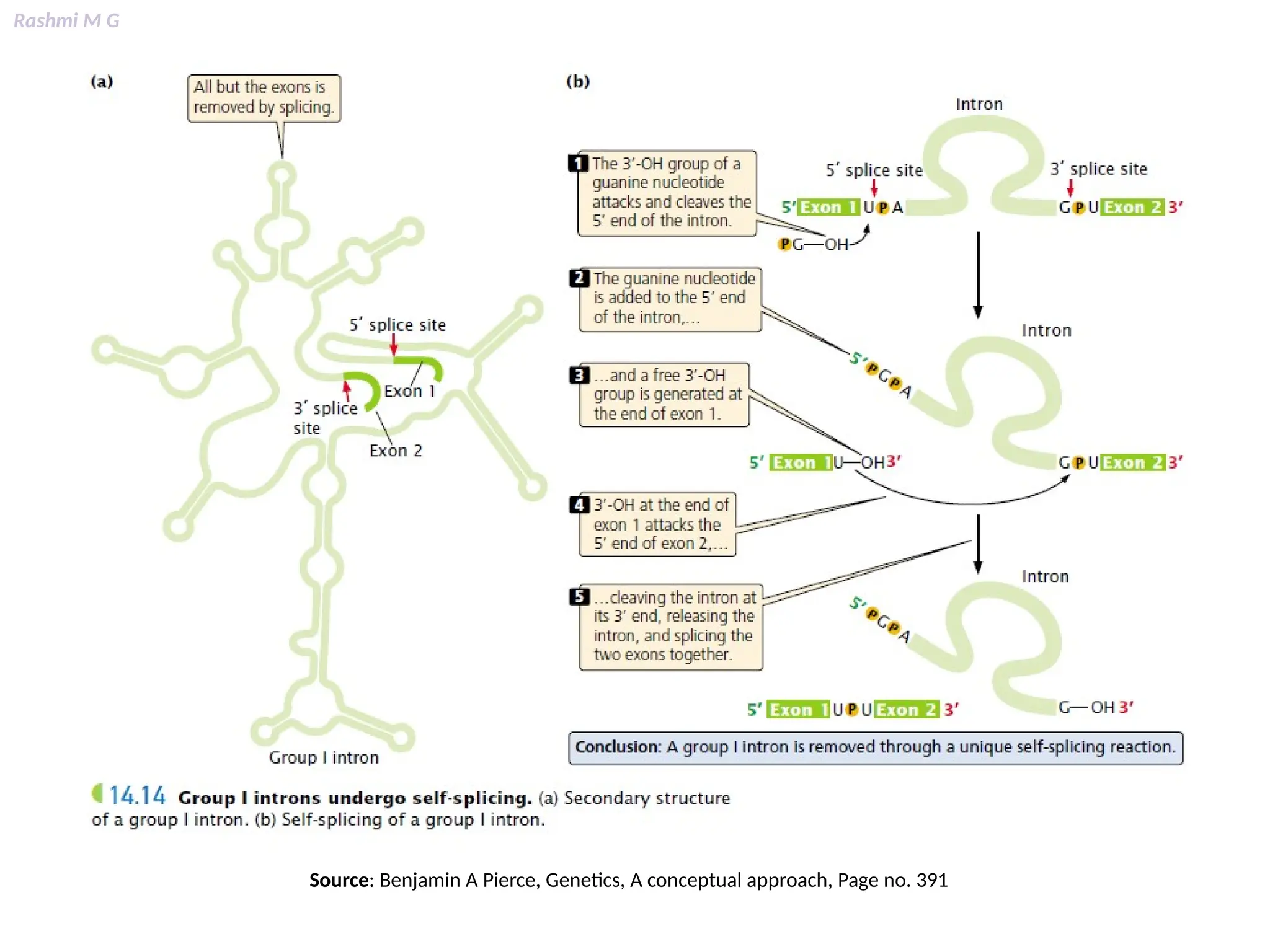







This PPT contains, Introduction Mechanisms of RNA processing Processing of pre-mRNA 5’ capping Polyadenylation Types of polyadenylation Intron splicing Exon and intron Definition Splicing apparatus Alternative splicing Trans splicing RNA editing Processing of pre-rRNA Group I and group II introns Processing of pre-tRNA mRNA degradation mRNA surveillance



































































